|
单细胞测序技术服务 靶向单细胞测序(lncRNA&mRNA) 单细胞测序 |
|
蛋白表达定量 DIA定量蛋白质组学 Label free非标定量 |
蛋白修饰定量 N-糖基化蛋白组学 O-GlcNAc修饰蛋白质组学 |
|
Ribo-seq Ribo seq(ribosome profiling) |
核糖体-新生肽链复合物(RNC) RNC联合 circRNA芯片 RNC联合 lncRNA芯片 RNC-seq |
|
NGS测序技术服务 环状DNA测序(eccDNA测序) |
PCR技术服务 环状DNA PCR技术服务 |
表观转录组学简介
表观转录组学(Epitranscriptomics)是近来兴起的热门领域之一,主要研究RNA所携带的化学修饰对基因表达的影响。生物体内的很多大分子都存在化学修饰,且都具有极其重要的作用。比如DNA或组蛋白上的化学修饰参与介导表观遗传调控,许多蛋白(尤其是酶类蛋白)的活性受到多种化学修饰的调节等等。迄今为止,在RNA上已发现了一百多种化学修饰。这些修饰大量分布在非编码RNA(ncRNA),特别是rRNA, tRNA和snRNA上,为ncRNA在翻译与剪接中发挥正常功能所必需。令人兴奋的是,研究人员发现m6A(N6-methyladenosine),m1A(N1-methyladenosine),m5C(5-methylcytidine),hm5C(5-hydroxylmethylcytidine),I(inosine)以及ψ(pseudouridine)等化学修饰也分布在真核生物mRNA上,影响mRNA的代谢与功能。特别是伴随着许多mRNA修饰酶(Writer)、去修饰酶(Eraser)和修饰识别蛋白(Reader)的新发现,mRNA化学修饰的可逆变化与动态调控重新激起了研究人员的兴趣,受到越来越多的关注[1]。
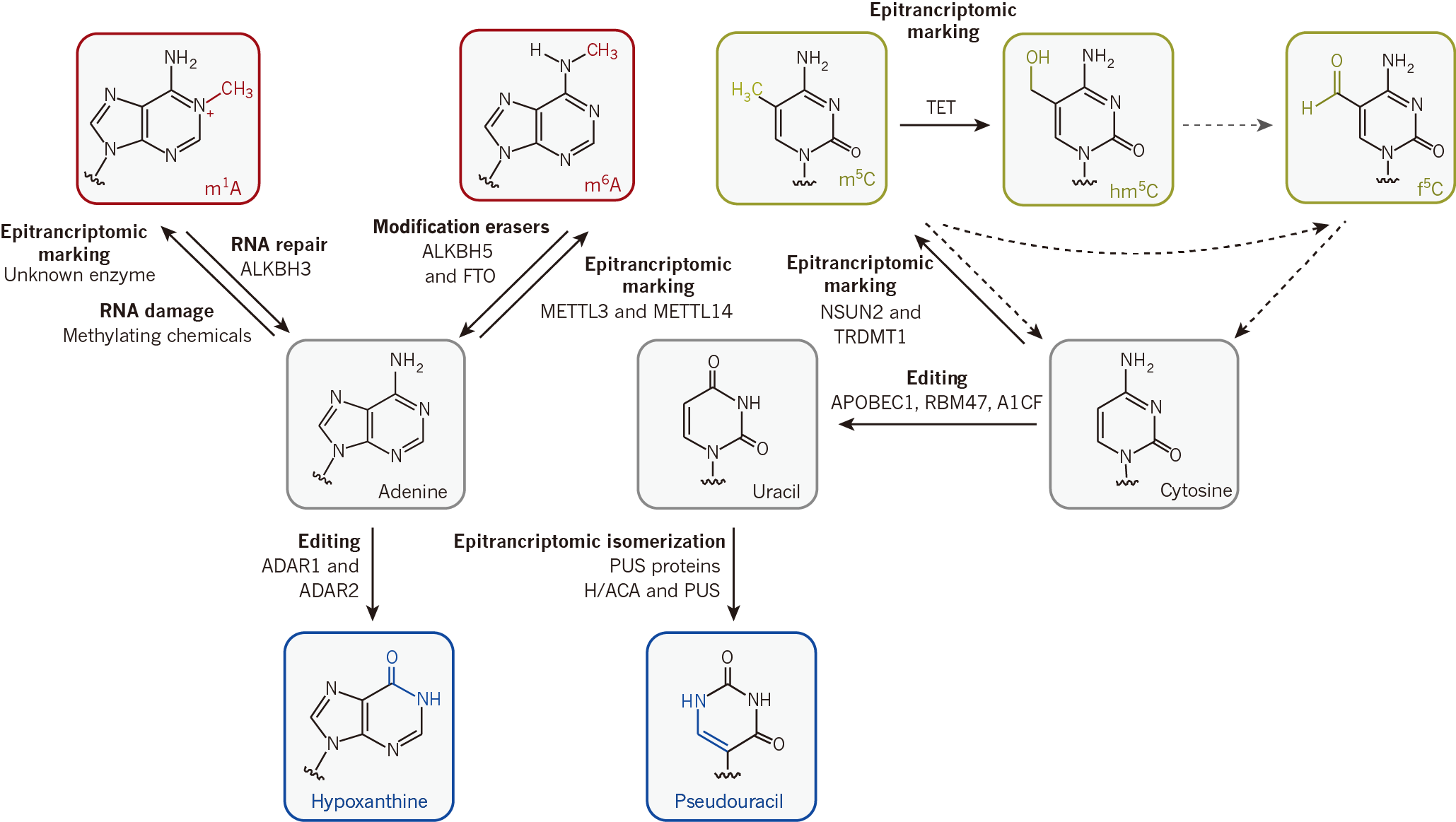
图1. 动态可逆的表观转录修饰途径[2]。
正如表观遗传修饰决定着基因何时何处转录一样,mRNA上存在的化学修饰可能代表着一种全新的转录后基因表达调控方式。尤其是动态可逆修饰的发现,意味着在mRNA的生命周期内,细胞可以通过添加或移除这些修饰,进行更直接、瞬时的基因表达调控,实现对外界环境变化的快速反应。我们知道,基因表达需要pre-mRNA加帽、可变剪接、多聚腺苷酸化以及将成熟mRNA转运到细胞质等过程的协调发生,细胞状态的改变也需要转录组精确受控的协同转变,生物节律的周期控制则需要协调更多更复杂层级的生物学过程。表观转录修饰可能是细胞实现这些协同过程的潜在方式之一。已有研究证明,m6A在细胞分化过程发挥不可或缺的作用[3]。另外,和肿瘤细胞中表观遗传修饰常发生错置一样,m6A在肿瘤、肥胖等疾病中也发生错置,且常与mRNA修饰酶(Writer)、去修饰酶(Eraser)和修饰识别蛋白(Reader)发生突变有关。
表1. 已发现的人属mRNA修饰酶(writer)、去修饰酶(Eraser)和修饰识别蛋白(Reader)。
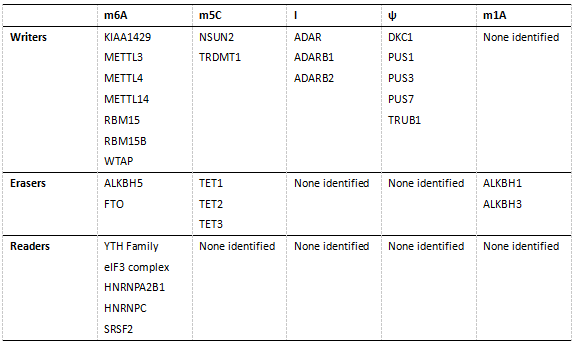
近几年来,表观转录组学研究取得了一系列突破性进展,然而其研究尚处于起步阶段。得益于检测技术的进步,表观转录修饰的研究步伐不断在加快。特别是高通量测序技术与质谱技术的发展,使得在表达丰度较低的mRNA中检测和鉴定化学修饰成为可能[6]。利用质谱技术(LC-MS/MS)检测总体mRNA中各修饰碱基的表达量,利用表观转录组测序技术(MeRIP-Seq)检测特异修饰在mRNA转录本上的丰度与分布,结合mRNA修饰酶(Writer)、去修饰酶(Eraser)和修饰识别蛋白(Reader)的敲除与过表达技术,全转录组测序技术,以及MeRIP-PCR技术,研究人员正在不断的开拓表观转录组学领域——扩展mRNA上存在的修饰种类,对转录本的修饰类型与位置进行注释,绘制不同物种间的保守修饰位点图谱,检测环境变化所引起的修饰表达与分布改变,阐明这些改变对mRNA生成、稳定性与翻译的影响,揭示动态修饰介导的生理病理学效应。
表观转录组学功能
目前对表观转录修饰影响细胞功能的作用机制仍知之甚少。这些化学修饰可能引起mRNA配对、热动力和折叠性能的改变,从而影响RNA的可变剪接、翻译、细胞定位、稳定性以及与蛋白的相互作用[2,7]。mRNA结构与功能的变化如何导致生物学功能的改变仍需要更进一步的深入研究。接下来,我们将从结构效应和生物学功能等方面对目前已知的mRNA表观转录修饰(m6A、m1A、m5C、hm5C、ψ、I)分别作简单介绍。
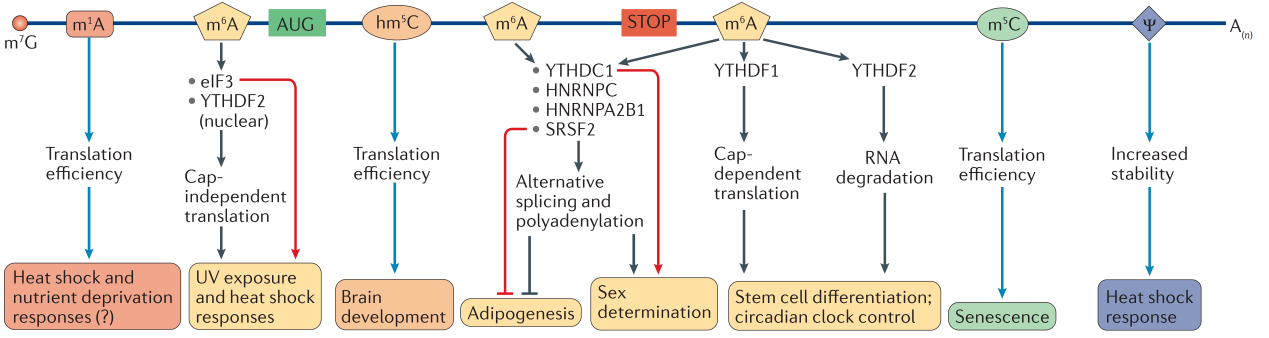
图2. mRNA修饰的生物学功能[7]。
m6A
m6A是真核生物mRNA上含量最丰富的化学修饰,由甲基转移酶复合物(包含METTL3,METTL14,WTAP,KIAA1429,RBM15,RBM15B)催化产生,可被去甲基化酶ALKBH5或FTO去除。目前已发现了多种特异性识别m6A位点的蛋白或复合物,包括YTH家族蛋白(YTHDF1-3,YTHDC1)、转录起始复合物eIF3、核糖核蛋白(HNRNPA2B1,HNRNPC)以及RNA结合蛋白SRSF2。m6A主要分布在终止密码子附近和3’UTR区,影响RNA配对、改变RNA二级结构、或被蛋白直接识别,进而调控mRNA的成熟、可变剪接、稳定性和翻译过程。与A-U配对相比,m6A-U配对较不稳定,引发RNA内部双链解旋与二级结构转变。m6A常在双链与单链的过渡区域堆叠,增强RNA转变后构象的稳定性。去甲基化则可以使mRNA恢复原来构象。这种构象转变可能导致mRNA与不同蛋白的相互作用改变,从而产生不同的生物学效应。m6A能直接被特定蛋白的疏水结构域所识别。比如,YTH家族蛋白可以特异性的识别m6A,特别是GGm6ACU保守序列。其成员之一YTHDC1识别并结合m6A,调控靶向mRNA的可变剪接。而另一成员YRHDF2与m6A结合后,招募CCR4-NOT复合物,促进靶向RNA的降解。在UV辐射或热休克反应中,转录起始复合物eIF3与5’UTR区的m6A结合,促进帽非依赖的翻译过程。编码区的m6A可被SRSF2识别,参与脂肪生成的调控。在果蝇中,YTHDC1同源蛋白识别性别致死mRNA上的m6A位点,调控其可变剪接,从而控制果蝇的性别。如前所述,m6A介导的mRNA稳定性调节也对干细胞分化与生物节律时钟控制十分重要。(图2) 此外,m6A也能通过影响mRNA与tRNA反密码子的配对速率与保真度,从而影响翻译延伸。
m1A
m1A是新近发现的可逆的表观转录修饰,能被RNA修复酶ALKBH3去除,目前尚未发现明确的m1A修饰酶与修饰识别蛋白。与m6A不同,m1A的表达丰度较低,主要分布在mRNA的5’UTR区,可能参与调节翻译起始过程。m1A能完全阻止Watson-Crick配对,引起RNA双链解旋,并促进RNA-蛋白的静电相互作用或RNA可变二级结构的形成。m1A的生物学功能仍属未知。有研究发现,在热休克或营养匮乏等压力条件下,细胞内的m1A表达水平上升,可能是通过促进帽依赖性翻译,参与细胞的应激反应。(图2)
m5C与hm5C
m5C广泛分布于tRNA与rRNA上,具有稳定tRNA二级结构、影响反密码子环构象、维持rRNA翻译保真等功能。新近的RNA测序结果发现,mRNA的编码区与非编码区上存在8000多个m5C位点,而且相当一部分位点集中在5’UTR与3’UTR区。m5C可由甲基转移酶NSUN2或TRDMT1催化形成,被双加氧酶TET氧化形成hm5C。hm5C可能经过进一步氧化形成f5C,进而变回胞嘧啶核苷(C)。m5C不影响碱基配对,但可能增强碱基堆叠以及RNA与蛋白的疏水作用。m5C具有多种生物学功能。p16 mRNA在被NSUN2酶添加m5C修饰后,其降解被抑制,稳定性增强。在细胞周期中,NSUN2的表达受到精密调控。NSUN2可在CDK1 3’UTR区添加m5C修饰,促进其翻译;同时在CDKN1B的5’UTR区添加m5C修饰,抑制CDKN1B的翻译。二者共同作用,增强细胞的增殖能力。m5C还与衰老相关基因的翻译控制有关,过表达NSUN2可以延缓复制性衰老的发生。作为m5C的氧化产物,hm5C也能增强翻译效率。hm5C在果蝇的脑中表达量很高,可能参与果蝇的脑部发育。(图2)
Pseudouridine (ψ)
假尿嘧啶核苷ψ,常被成为第五类核苷酸,由尿嘧啶核苷(U)异构化形成。在人的细胞和小鼠组织的mRNA中,ψ/U的比率约为0.2-0.6%。尿嘧啶与假尿嘧啶的异构化反应由PUS酶单独,或与H/ACA核糖核蛋白一起,催化完成。假尿嘧啶能减少RNA构象的可变性,增强碱基配对稳定性以及与蛋白之间的的极性相互作用。假尿嘧啶可能调控mRNA稳定性和基因表达,参与酵母的热休克反应,但具体机制目前尚不明确。(图2)
Inosine (I)
肌苷修饰,常称为A-to-I编辑,是高等真核生物里最常见的一种RNA编辑方式,由腺苷酸脱氨酶ADAR完成。A-to-I编辑主要发生在非编码区或内含子区的Alu元素内。A-to-I编辑完全改变了碱基配对特性,AU配对转变为IC配对,从而改变所编码的氨基酸。比如,A-to-I编辑将大脑谷氨酸受体中的谷氨酰胺重编码为精氨酸,导致了钙离子通透性的改变。此外,A-to-I编辑还具有改变可变剪接、调节miRNA的产生与功能、以及监控先天性免疫反应等作用。
表观转录组学研究方法
康成生物推出了多层次、全方位的表观转录组学研究工具供客户选择,包括:
m6A单碱基分辨率芯片技术服务(在单碱基水平上定量和定位 RNA m6A修饰位点,是m6A机制研究利器)
mRNA&lncRNA 表观转录组芯片(m6A)技术服务(在转录本水平上定量检测m6A修饰百分比及变化,适用于mRNA, lncRNA, pre-miRNA, pri-miRNA, snoRNA, 和 snRNA)
circRNA 表观转录组芯片(m6A)技术服务(在转录本水平上定量检测检测m6A修饰百分比及修饰变化,适用于环状RNA)
mRNA甲基化测序(m6A修饰位点与分布检测)
MeRIP-PCR (携带m6A修饰的特定转录本检测与验证)等。
LC-MS mRNA碱基修饰检测(mRNA上各类修饰总量检测)
LC-MS tRNA碱基修饰检测(tRNA上各类修饰总量检测)
NuRNA™ Epitranscriptomics PCR Array(修饰添加、去除与识别蛋白基因表达水平检测)
NuRNA™ tRNA Modification Enzymes PCR Array(检测tRNA修饰酶及相关蛋白因子)
这些工具的组合使用,可以帮助科研人员从不同视角、不同层面研究表观转录修饰的特性与功能,探寻修饰异常与疾病的联系,为疾病治疗提供新的思路。


