|
单细胞测序技术服务 靶向单细胞测序(lncRNA&mRNA) 单细胞测序 |
|
蛋白表达定量 DIA定量蛋白质组学 Label free非标定量 |
蛋白修饰定量 N-糖基化蛋白组学 O-GlcNAc修饰蛋白质组学 |
|
Ribo-seq Ribo seq(ribosome profiling) |
核糖体-新生肽链复合物(RNC) RNC联合 circRNA芯片 RNC联合 lncRNA芯片 RNC-seq |
|
NGS测序技术服务 环状DNA测序(eccDNA测序) |
PCR技术服务 环状DNA PCR技术服务 |
DoGs在细胞衰老中介导转录干扰
DoGs可以作为反义RNA来调控蛋白编码基因的表达。在衰老细胞中,衰老触发了反义通读RNA(START RNA)的产生,即DoGs[1]。START RNA抑制其相应正义RNA的表达。
另一个案例是位于拟南芥基因组的冷敏感区域的SVK基因。 SVK的RNAPII介导的通读转录导致产生了一个lncRNA- asCBF1,该lncRNA在CBF1的反义链上,与CBF1位置重叠[2]。 由于SVK-asCBF1级联转录的RNAPII与CBF1的RNAPII转录碰撞,导致CBF1的转录抑制(图1)。这种SVK-asCBF1级联转录的机制严格控制了冷胁迫条件下CBF1的表达。
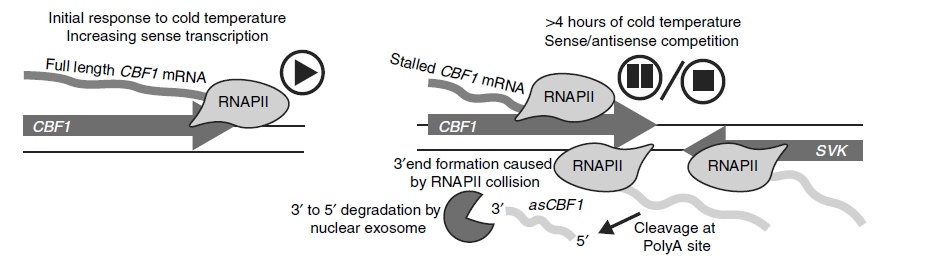
图1:通过反义DoG RNA控制CBF1编码基因的表达。 (左图)在正常温度下,CBF1可以顺利转录。 (右图)在暴露于寒冷环境后,CBF1的反义方向上SVK转录增加。 SVK的转录通读导致与CBF1的3’端反义的lncRNA asCBF1转录。 在两条链上转录RNAPII的占据增加导致RNAPII的碰撞和CBF1正向转录的停顿,从而导致CBF1完整mRNA产量减少[2]。
在病毒感染中DoG调控基因组的三维结构
在流感病毒(IAV)感染期间,病毒非结构蛋白NS1诱导高活性宿主基因的通读转录,导致凝聚素从宿主染色质中解离,染色质环消失,以及在通读转录区域中染色质的解聚(图2)[3]。
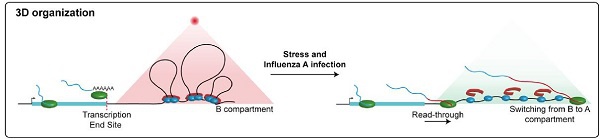
图2:通读转录改变染色质结构。流感病毒(IAV)蛋白NS1干扰转录终止,使RNAPII在终止位点之后继续前进,导致染色质环的消失和局部染色质解聚 [3]。
相关服务
Downstream-of-Gene Transcript (DoG RNA)芯片
相关资源
什么是基因下游(DoG)RNA?
DoGs形成了嵌合RNA和环状RNA
参考文献
1. Muniz L et al: Control of Gene Expression in Senescence through Transcriptional Read-Through of Convergent Protein-Coding Genes. Cell Rep 2017, 21(9):2433-2446.[PMID: 29186682]
2. Kindgren P, Ard R, Ivanov M, Marquardt S: Transcriptional read-through of the long non-coding RNA SVALKA governs plant cold acclimation. Nat Commun 2018, 9(1):4561.[PMID: 30385760]
3. Heinz S et al: Transcription Elongation Can Affect Genome 3D Structure. Cell 2018, 174(6):1522-1536 e1522.[PMID: 30146161]


