|
单细胞测序技术服务 靶向单细胞测序(lncRNA&mRNA) 单细胞测序 |
|
蛋白表达定量 DIA定量蛋白质组学 Label free非标定量 |
蛋白修饰定量 N-糖基化蛋白组学 O-GlcNAc修饰蛋白质组学 |
|
Ribo-seq Ribo seq(ribosome profiling) |
核糖体-新生肽链复合物(RNC) RNC联合 circRNA芯片 RNC联合 lncRNA芯片 RNC-seq |
|
NGS测序技术服务 环状DNA测序(eccDNA测序) |
PCR技术服务 环状DNA PCR技术服务 |
长链非编码RNA是一类在细胞活动中发挥重要作用的调控型RNA,但这些RNA在发育过程中的功能却有很多未知的地方。本研究,作者应用Arraystar Mouse lncRNA芯片发现了一系列此前未被发现过的成簇的、在大脑非印记区域的保守的lncRNA(CNIB,conserved nonimprinting lncRNA clusters in brain)。在发现的7个lncRNA簇中,人的CNIB1 位于Chr 9q33.3,且在斑马鱼到人都有保守性。Chr 9q33.3-9q34.11微缺失与人指甲-髌骨综合征(伴随有发育和视觉缺陷)有关联。通过在斑马鱼上构建CNIB1 缺失等位基因模型,作者证明了CNIB1 对正常生长、发育及视觉活动是必要的。最后,作者发现CNIB1 对视觉功能的影响是通过一个眼睛发育的调控因子Lmx1bb 来发挥调节作用的。综合以上,作者的研究表明CNIB1 lncRNA簇对斑马鱼的发育是非常重要的,同时为人类Chr 9q33.3-9q34.11微缺失综合征提供了一个lncRNA簇调节的病理生理学机制。该研究发表在Nucleic Acids Research(IF:11.147)。(芯片实验由康成生物提供技术服务)
研究背景
一个基因簇是由一组紧密分布在同一条染色体并协调表达或有相关功能的基因构成的。基因簇的发现加深了我们对于基因组功能和进化的理解。近些年,一些小的非编码RNA簇已经被报道,如Sfmbt2 miRNA 簇及snoRNA簇。长链非编码RNA(lncRNA)是一大群超过200nt且在RNA水平调控多种细胞活动的RNA。一些独立的研究发现,lncRNA也可以形成簇。
人的大脑是最丰富表达lncRNA的器官之一,被报道表达有20000个lncRNA,该数量与人全部基因组的编码基因数目相当。功能研究发现,lncRNA与神经细胞的类型决定、可塑性、突出传递、高级认知能力及大脑发育相关。尽管lncRNA的数量也在增加,但是中枢神经系统中表达的lncRNA的图谱也仅仅才开始被理解。
斑马鱼是一种良好的研究人类疾病的脊椎动物模式系统,应用斑马鱼进行的研究在发育生物学、神经生物蛋白编码基因进化理论等多个领域产生过前沿成果。与哺乳动物类似,斑马鱼的lncRNA也被报道在物种间的保守性低。保守的lncRNA仅占斑马鱼lncRNA的 9%不到,467个lncRNA与灵长类和啮齿类动物是直系同源的。有假设认为,尽管大部分lncRNA在复杂的生理学活动中作为调控因子发挥作用,仍有小部分lncRNA是对胚胎发育、生存和发育必不可少的。因此,利用斑马鱼这种模式生物开展保守的lncRNA研究,能帮我们进一步理解人类疾病相关的lncRNA的生物学功能和分子作用。
本文,作者发现了一个新的被命名为CNIB s 的lncRNA基因簇。在7个CNIB 中,CNIB1 是从斑马鱼到灵长类保守度最高的一个,并且与人类的Chr 9q33.3-9q34.11微缺失综合征有密切关系。结果表明,CNIB1 座位对于斑马鱼的正常发育、运动能力及视觉功能是必要的。CNIB1的缺失也被发现存在于一部分Chr 9q33.3-9q34.11微缺失综合征病人中。
技术路线
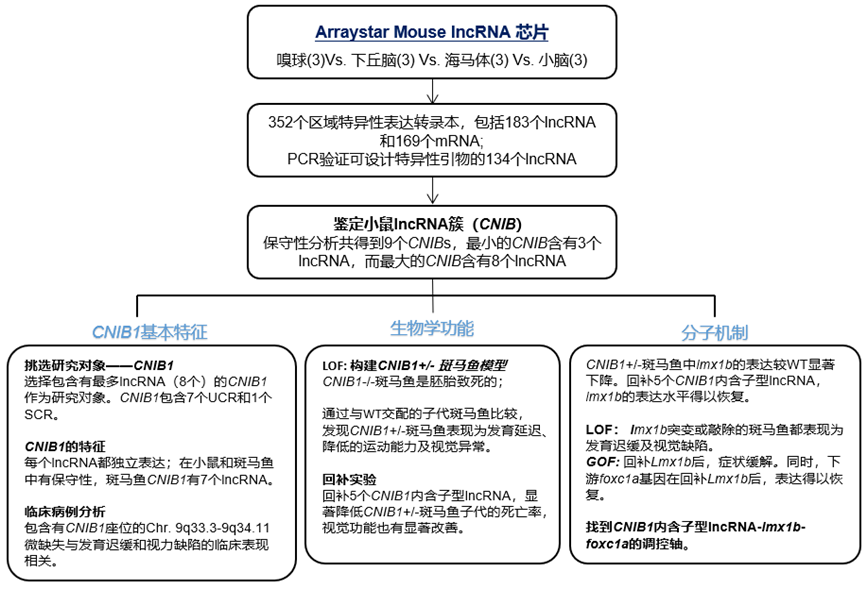
研究结果
分析大脑中高表达的lncRNA
为了研究大脑不同部位的lncRNA表达谱,作者应用Arraystar Mouse lncRNA芯片检测了成年雄性小鼠大脑中的嗅球、下丘脑、海马体和小脑中的lncRNA表达谱。在四种脑部位中,共筛选到17619表达的mRNA和20202个表达的lncRNA。为了鉴定不同大脑区域专一性表达的转录本,作者设置了更严格的标准,共鉴定到了352个区域特异性表达转录本,其中包括183个lncRNA和169个mRNA。除了49个lncRNA因无法设计特异性引物外,作者将其余134个转录本都进行了验证。
鉴定保守的、位于非印记区且在大脑中表达的lncRNA簇
在鉴定大脑高表达的lncRNA时,有34个lncRNA座位是紧挨着的,形成位于7条染色体的9个簇,其中最小的簇,每个都含有3个lncRNA。作者通过分析已发表的野生型小鼠的RNA seq数据确认了这34个lncRNA的表达。通过对这9个lncRNA簇的保守性分析,有7个被认为是保守的,因为这些lncRNA要么来源于超保守元件(UCR)或SCR(shared conserved patches)。这7个保守的簇被命名为CNIB lncRNA簇(conserved nonimprinting lncRNA clusters in brain),详见下表:
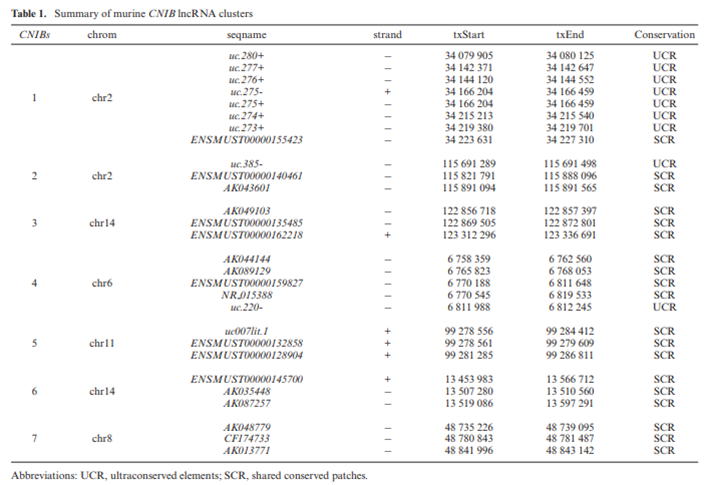
CNIB1 lncRNA的特征
作者从7个CNIB 中选择了有最多lncRNA的CNIB1 作为研究对象。在CNIB1 的8个lncRNA中,有7个UCR(uc.280+, uc.277+, uc.276+,uc.275+, uc.275-, uc.274+和 uc.273+) 和一个SCR(ENSMUST00000155423),除了uc.274+,其余6个UCR和1个SCR在斑马鱼中都存在。DNA序列水平分析发现,uc.280+有在进化中76%的相似性,而其他7个CNIB1 lncRNA则有几百的、在进化中相似性达87%-100%的序列。此外,这些CNIB1 lncRNA在基因位置及总分布(相同的合成顺序、链)上也是保守的。
作者通过已发表的ChIP-seq数据研究了CNIB1 lncRNA的特征,有如下发现:
1)每个lncRNA都有唯一的转录本起始位点,且是独立被转录的;
2)小鼠CNIB1 lncRNA都有5’端帽子;
3)大部分CNIB1 lncRNA在小鼠和斑马鱼中都有多个相似异构体。对5个可检测的CNIB1 lncRNA(uc.277+, uc.276+,uc.275+, uc.275-,和uc.273+)的PCR结果发现,在小鼠和斑马鱼中的表达模式是相似的。这其中,有几个CNIB1 lncRNA在小鼠和斑马鱼的大脑及眼睛里的表达升高。在斑马鱼幼体上,Whole-mount ISF结果也进一步确认了这一结果。
综合以上,CNIB1 lncRNA在小鼠和斑马鱼上有相似的特征及表达模式,表明CNIB1在进化中有保守的功能。
包含有CNIB1座位的Chr 9q33.3-9q34.11微缺失与发育迟缓和视力缺陷的临床表现相关
人同源的CNIB1序列位于Chr 9q33.3。而人类Chr 9q33.3-9q34.11微缺失指甲-髌骨综合征(NPS)有关。NPS是一种常染色体显性遗传病,症状为指甲发育不良和髌骨发育不全或发育不良。因其复杂的遗传背景,个体的患病程度不一。同时,包括肾以及视觉系统也可能会受到影响。LMX1B ,位于Chr 9q33.3,是LIM同源异型转录因子成员,当前研究认为其与NPS的表型相关。然而,尚不清楚CNIB1 区域是否参与NPS的发展。
截止当前,10例有完全的临床和遗传学特征的NPS患者已被报道。下图B的b区域圈出了共同的Lmx1b 缺失区域。在这10个患者中,3个患者都有CNIB1 缺失,而这3个患者都有严重的发育迟缓和神经系统表现。 另外,有2个患者有视觉缺陷。这个信息提示我们,视觉异常和NPS的其他症状或归咎于CNIB1区域。
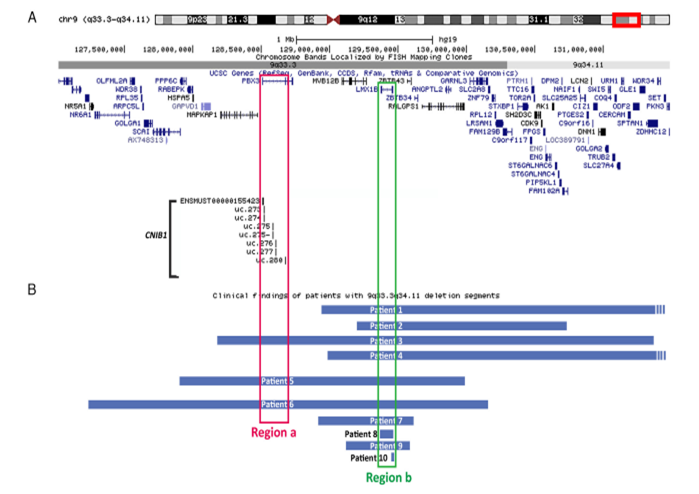
有趣的是,除了CNIB1 座位与NPS症状的相关性外,作者发现,其他3个CNIB(CNIB2、CNIB3和CNIB4)也与人的中枢神经系统缺陷有关。CNIB2 是从Meis2 基因转录而来。在17个有中枢神经系统缺陷并带有Chr 15q14微缺失的患者中,最小的共有关键片段即是CNIB2 和它邻近Meis2 的区域。CNIB3 也是在超过一半的有Chr 13q32.2-33.2微缺失患者(表现为严重的神经管缺失,眼/手/脚异常及面部先天性畸形)中的最小共有片段。Dlx-5,Dlx-6 是CNIB4 的宿主基因,被报道是神经胶质命运和少突胶质细胞系成熟的关键调控因子。NR 015388 和 ENSTMUST00000159827 (也叫做Evf2 )都在CNIB4 座位内。这两个lncRNA能与BRG1 相互作用,介导染色质重塑。尽管未在CNIB4 lncRNA上发现基因组突变,但我们发现BRG1蛋白的RNA结合结构域的突变与Coffin-Siris Syndrome(一种神经发育异常疾病)相关。
CNIB1+/-斑马鱼表现为发育延迟、降低的运动能力及视觉异常
为了研究CNIB1在体内的功能,作者用CRISPR-Cas9系统构建了CNIB1突变等位基因斑马鱼。斑马鱼CNIB1包含有7个lncRNA:ENSMUST00000155423 , uc.273+, uc.275+, uc.275-, uc.276+,uc.277+和uc.280+(以染色体正义链5’-3’顺序列出)。其中uc.280+和ENSMUST00000155423与邻近的pbx3b基因的外显子有重叠,其余5个转录本都连续分布在pbx3b旁侧的内含子上,远离pbx3b的剪接位点,且在该区域无其他已知或被注释的RNA。为了避免pbx3b的影响,作者设计了sgRNA,敲除了这5个lncRNA对应的内含子区域,得到的斑马鱼称为CNIB1 KO斑马鱼。通过基因分型检测,在F1代共得到两个独立的敲除等位基因的斑马鱼,CNIB1+/- 1共有48247bp的缺失,CNIB1+/- 2共有48221bp的缺失。这些同源异型的F1代再交配产生F2代。有趣的是,F2代中没有纯合子后代,而WT和杂合子的比值为1:2。后续的研究发现,尽管这两组在刚受精及受精4h和12h后的受精卵数量是几乎相同的,但在受精24h后,杂合子杂交存活的鱼卵数降为WT杂交鱼卵数的3/4。这些结果提示我们CNIB1-/- 斑马鱼是胚胎致死的,或在细胞分裂时已开始死亡。
在CNIB1杂合子斑马鱼中,与WT斑马鱼相比,所有的5个参与的lncRNA都显著下调了。相反,CNIB1和pbx3b的两个邻近基因gapvd1和mvb12bb 则无显著变化。为了排除是CNIB1+/- 影响了pbx3b 基因的剪接,作者对pbx3b 的2号外显子和3号外显子的结合区进行测序,并设计多个PCR引物来检测pbx3b 的可变剪接事件。作者的结果显示,在WT和CNIB1+/- 中,2号外显子和3号外显子的结合区域以及pbx3b 基因的可变剪接保持一致。这些结果表明,CNIB1+/- 对宿主基因pbx3b 的剪接没有影响。
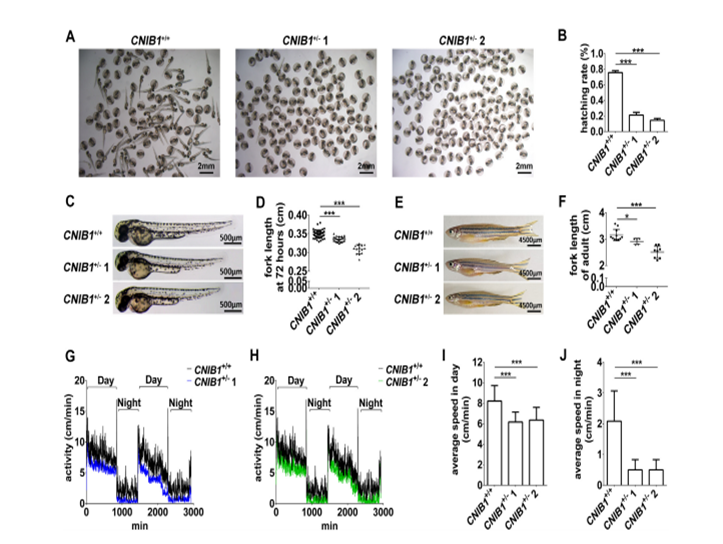
此外,作者发现,F2 CNIB1+/- 斑马鱼与WT间交配产生的后代相比,有发育迟缓、降低的运动能力及视觉异常:
1) 在受精48h后,超过75%的WT间交配产生的受精卵孵化了,而CNIB1+/- 1或CNIB1+/- 2交配后的卵孵化的比例则小于20%(下图A&B);
2) 受精72h后及4个月的成年的斑马鱼,CNIB1+/- 1或CNIB1+/- 2交配后的后代的体长都明显更短,在F1代中也有类似的结果(下图C、D、E&F);
3) 运动能力检测发现,CNIB1+/- 1或CNIB1+/- 2交配的后代的白昼和黑夜的平均运动速度都显著下降(下图G-J)。
4) CNIB1+/- 1或CNIB1+/- 2交配的后代的眼睛更小(下图A&B)且视动力反应(OKR)检测到的降低的gain value提示有眼速障碍(下图C&D)。
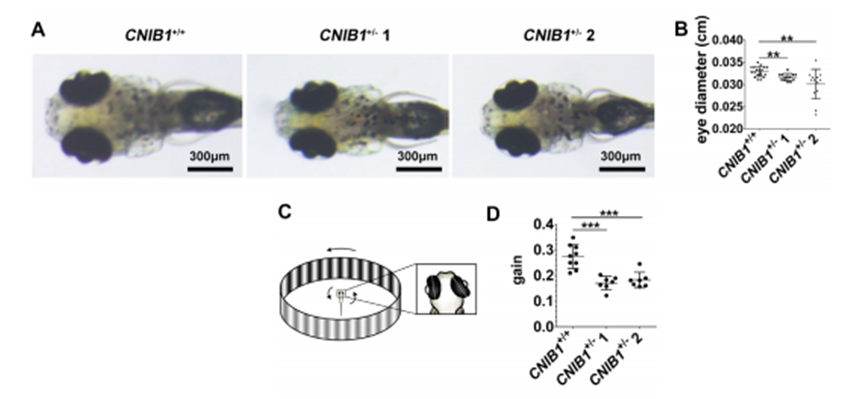
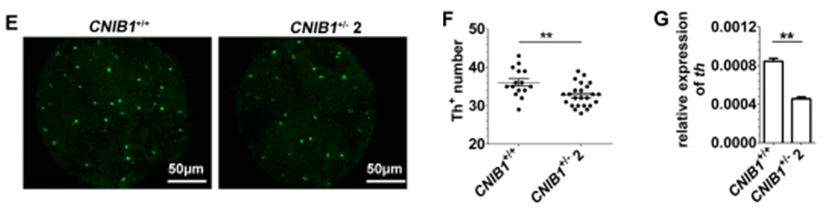
为了进一步研究CNIB1对眼睛发育和功能的影响,我们检测了CNIB1敲除斑马鱼的视觉系统的结构是否受到影响。通过对眼睛的DAPI染色,结果显示,在CNIB1+/- 2和WT斑马鱼的视网膜上,并未有总体的形态差异。这些结果提示我们,CNIB1+/- 2的缺陷并非是结构上的。视网膜多巴胺是多巴胺神经元在酪氨酸羟化酶作用下产生的,在光适应、细胞生存和眼睛生长等方面发挥多种和营养相关的作用。因此,作者检测了视网膜多巴胺神经元的数量以及酪氨酸羟化酶基因的表达水平。作者的结果显示,与WT斑马鱼相比,CNIB1+/- 斑马鱼视网膜上酪氨酸羟化酶标记的多巴胺神经元数量显著下降(下图E&F)。同时,CNIB1+/- 2斑马鱼的酪氨酸羟化酶mRNA的水平也显著降低了(下图G)。
综合以上结果表明,CNIB1座位对斑马鱼的正常发育、运动能力和视觉功能都是必需的。
在斑马鱼中,CNIB1内含子型lncRNA是通过lmx16b基因调节视觉功能的
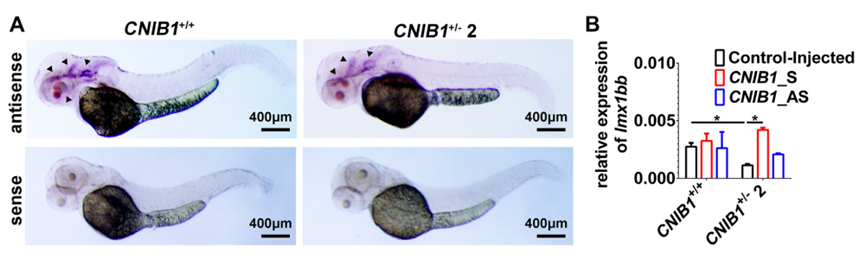
为了确定是否是因为CNIB1 lncRNA导致了杂合子交配后代上发现的视觉缺陷,作者在体外WT间或CNIB1+/- 2间交配的胚胎中注射入5个CNIB1内含子型lncRNA的混合物(CNIB1_S),以注入无RNA(Control-Injected)及一个uc.273+,uc.276+,uc.277+的反义转录本(CNIB1_AS)作为阴性对照。作者的RT-PCR结果显示,在受精48h后的胚胎中检测到RNA的高表达。进一步研究发现,与(Control-Injected)或CNIB1_AS相比,注射了CNIB1_S 的CNIB1+/-2交配的F2代斑马鱼的死亡率更低,且OKR的数据得到显著改善。作者的结果表明CNIB1内含子型的RNA在视觉功能中发挥关键作用。但实验并未排除整个CNIB1座位也可能参与其中。
Lmx1b,在斑马鱼中是lmx1bb 或lmx1b.1,是chr9q33.3-9q34.1微缺失综合征的可能治病基因,被报道在斑马鱼眼睛发育中影响fgf 调节的视网膜patterning。为了研究可能的lmx1bb 参与CNIB1介导的视觉调控,作者通过原位杂交检测了CNIB1是否调控lmx1bb 的分布及表达。结果显示,与WT斑马鱼相比,在受精48h后,lmx1bb 在CNIB1+/-2斑马鱼的眼睛和大脑中的表达水平显著下降了(下图A&B)。此外,在过表达CNIB1 lncRNA(CNIB1_S)后,lmx1bb的水平可恢复至正常水平(下图B)。
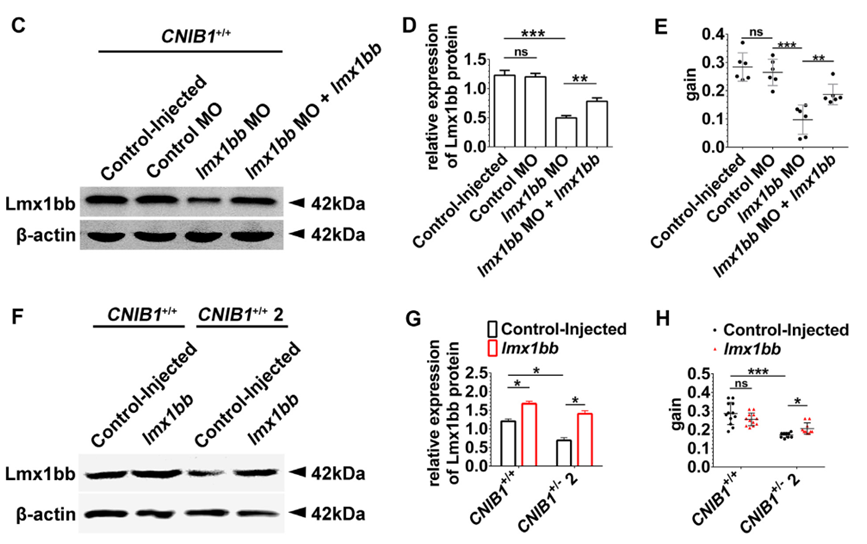
有报道称,lmx1bb 突变斑马鱼和lmx1bb morphants 都有眼睛发育迟缓及视觉缺陷。与已公开的结果相似,作者的结果也显示,lmx1bb morphants也有视觉障碍,而这可通过共注射lmx1bb mRNA得到挽救(下图C-E)。尽管过表达很多转录因子会导致WT胚胎的异常,有报道,在蛙、鸡和小鼠中过表达lmx1bb 不会发生发育和视觉异常。同样,在斑马鱼中过表达lmx1bb 也未产生明显异常。此外,与CNIB1+/+间交配的后代相比,在CNIB1+/-2间交配的后代中过表达lmx1bb会改善它们的视觉功能(下图F-H)。小鼠中lmx1bb 的下游靶基因foxc1a\eya2和pax8,在CNIB1+/-2斑马鱼中也下调了。然而在斑马鱼中过表达lmx1bb,仅有foxc1a 的表达恢复。
综合以上结果表明,CNIB1的活性至少部分是通过CNIB1内含子型lncRNA-lmx1bb-foxcta这个调控轴来实现的。
研究总结
1. 作者使用Arraystar Mouse lncRNA芯片发现了多个、来源于大脑非印记区的lncRNA簇,并命名为CNIB。并重点研究了一个在斑马鱼到小鼠上高保守的CNIB1作为研究对象。
2. CNIB1与宿主基因pbx3在功能和表达上是完全独立的
3. 从斑马鱼到人类,CNIB1都是和lmx1b位于同一条染色体上,lmx1b基因在小鼠和斑马鱼中对于眼睛发育和视觉功能都是必需的,且CNIB1调控下游的lmx1b及foxc1a。
4. CNIB1+/-的后代及lmx1b突变的动物都有发育迟缓和视觉功能异常,而通过外源补充lmx1bb,CNIB1+/-的视觉功能得以恢复。
综合以上,作者的研究提示了在发育中一个保守的CNIB1-lmx1b的功能,同时也提示了Chr. 9q33.3-9q34.11微缺失综合征中有CNIB1的参与。此外,lncRNA簇或许也存在于其他体系中,这些lncRNA簇的发现,将加深我们对lncRNA在进化和人类疾病中的作用的理解。


